

Marine ecological models used in EMoSEM
MIRO&CO-3D
MIRO&CO, a biogeochemical model dedicated to the study of eutrophication, results from the coupling between the COHERENS-3D hydrodynamic model (Luyten et al., 1999) and the biogeochemical model MIRO (Lancelot et al., 2005). It has been implemented in the Channel and Southern Bight of the North Sea (Lacroix et al. 2007) with realistic forcing (meteorological conditions and river loads) for the period 1991–2006. The model simulates the annual cycle of inorganic and organic carbon and nutrients (nitrogen, phosphorus and silica), phytoplankton (diatoms, nanoflagellates and Phaeocystis), bacteria and zooplankton (microzooplankton and copepods).
As a consequence of eutrophication, the spring bloom in the Belgian Continental Shelf (BCS) is dominated almost every year by the nuisance colonial species Phaeocystis. The nuisance of Phaeocystis bloom is usually measured by the intensity of its spring maximum and by the duration of the bloom. It may also be estimated with the annual integral of excessive abundance, IEB (Fig.1).
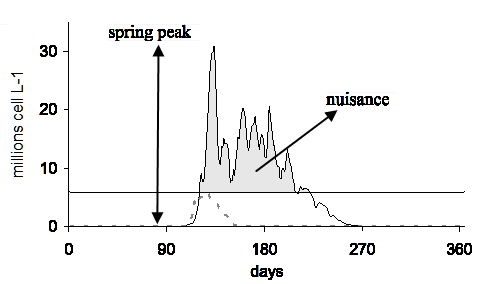
Figure 1. Modeled time series of Phaeocystis colonies abundance in the Belgian Coastal Shelf for an oligotrophic station (dotted line), and a coastal eutrophicated station (black line). The horizontal line indicates the abundance above which colonies become inedible for copepods (6 millions cells L-1). The grey area is a measure of Phaeocystis ecological nuisance, the integral of excessive abundance (IEB).
The spatial distribution of the Phaeocystis spring maximum illustrates the role of the rivers in marine eutrophication, as the maximum values are located close to the estuaries and along the coastal zones (Fig. 2). The spatial distribution of Phaeocystis nuisance also shows that, in areas further from the rivers influence, Phaeocystis may in some locations never reach problematic abundances.
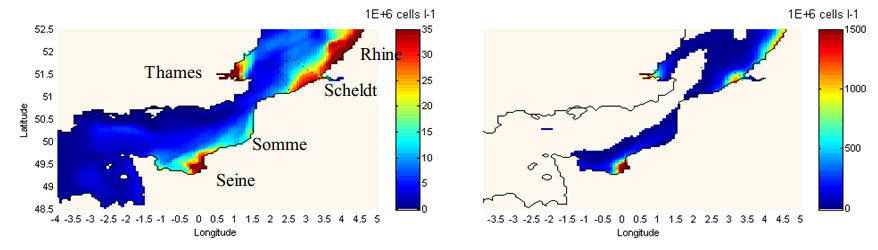
Figure 2. 3D-MIRO&CO model results in the English Channel and the Southern Bight of the North Sea for the year 2000. Left: map of Phaeocystis spring maximum abundance (not synoptic). Right: annual map of Phaeocystis IEB (see Fig.1). The white area depicts the area where Phaeocystis abundance never exceeds 6 millions cells L-1 during the year. (Source)
Additional information: see MUMM Ecosystem models, MUMM-REMSEM and AMORE.
ECO-MARS3D
The ECO-MARS3D model is currently a component of the French PREVIMER project of Coastal Operational Oceanography (previmer.org), and is based on the MARS3D hydrodynamical code (Lazure and Dumas, 2008). The application to the French Atlantic shelf is based on a regular grid with 4x4 km meshes and 30 sigma levels, which covers the Bay of Biscay, the English Channel and the southern part of the North Sea, up to the Rhine estuary; it extends from 8.13°W to 5.0°E, and from 43.17°N to 52.75°N (Fig. 3). It is an extended and re-calibrated version of a first model of the same space resolution, but limited to the Bay of Biscay proper, which was built as a Previmer demonstrator. This has been run operationally during five 6 years (2007-2012) on the previmer.org site, and used also for a long off-line run (1972-2008) for an environmental approach of pelagic fish fluctuations (Huret et al., 2013).
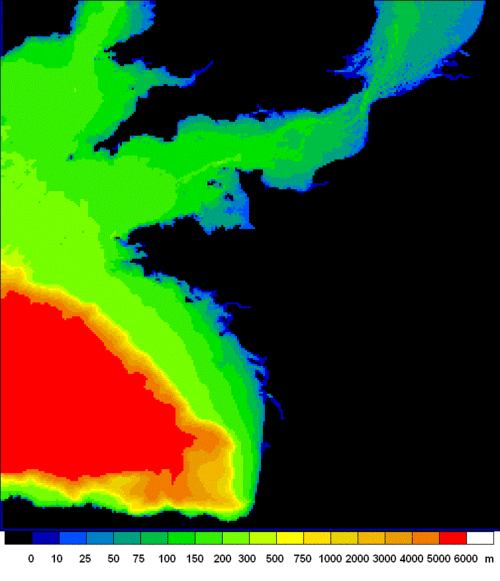
Figure 3. Bathymetry of the ECO-MARS3D model
Mechanical forcing is made by barotropic sea-level oscillation at the oceanic boundaries (provided by a 2D model covering the whole North-East Atlantic), and wind and atmospheric pressure at the sea surface; these are provided by the Arpege model of Météo-France with a 30 km and 6 h space-time resolution. Measured daily discharges as well as monthly river temperatures are provided on line by the Seine-Normandy, Loire–Brittany and Adour–Garonne River Basin Agencies, for the 5 main French rivers: Adour, Gironde, Loire, Vilaine, Seine. For all the other rivers in the domain, only discrete measurements of flow rates made in recent years are available for validation; in operational mode, the daily flow rate of these rivers is deduced from the measured flow rate of the nearest main river by linear regression. River daily concentrations for inorganic and organic dissolved nutrients are computed from empirical statistical relationships involving flow rate and time fitted to historical data (Guillaud and Bouriel, 2007). Suspended particulate matter is set to the maximum of ambient climatological monthly mean distribution derived from satellite data (Gohin et al., 2005) and the suspended matter brought by the rivers, which is simply simulated as a particulate conservative tracer, with uniform and constant settling velocity. At the open boundaries, all the biogeochemical state variables are imposed following a zero gradient condition.
The basic biogeochemical model contains 17 state variables, describing the nitrogen, phosphorus and silicon cycles and the dissolved oxygen in the pelagic ecosystem. Three limiting dissolved inorganic nutrients are considered: nitrogen, with nitrate and ammonium separately, phosphorus, and silicon. Phytoplankton is divided into 3 groups: diatoms, dinoflagellates and nanoflagellates, with concentrations expressed in nitrogen currency (Dussauze, 2011). Chlorophyll (Fig. 4) is deduced from the nitrogenous state variables of the model by an empirical Chl/N ratio, computed as a Smith-like formula depending on the local extinction coefficient. There are two zooplanktonic components, expressed in nitrogen units: the microzooplankton, which eats nanoflagellates and detrital particulate matter everywhere, along with diatoms in oceanic regions (depth>200m), and the mesozooplankton, which eats diatoms, dinoflagellates and microzooplankton. So, in this model, diatoms do sink, whereas nanoflagellates and dinoflagellates do not (they are considered as able to maintain at any depth in calm water, thanks to motility). Three particulate detrital variables (detrital N, detrital P, detrital Si) close the biogeochemical cycles, and settle in the water column; in the bottom layer, each settling fraction is partially transferred to a fixed state variable, which can give back to the water layer some particulate material through erosion by currents, and some dissolved equivalent after remineralisation.
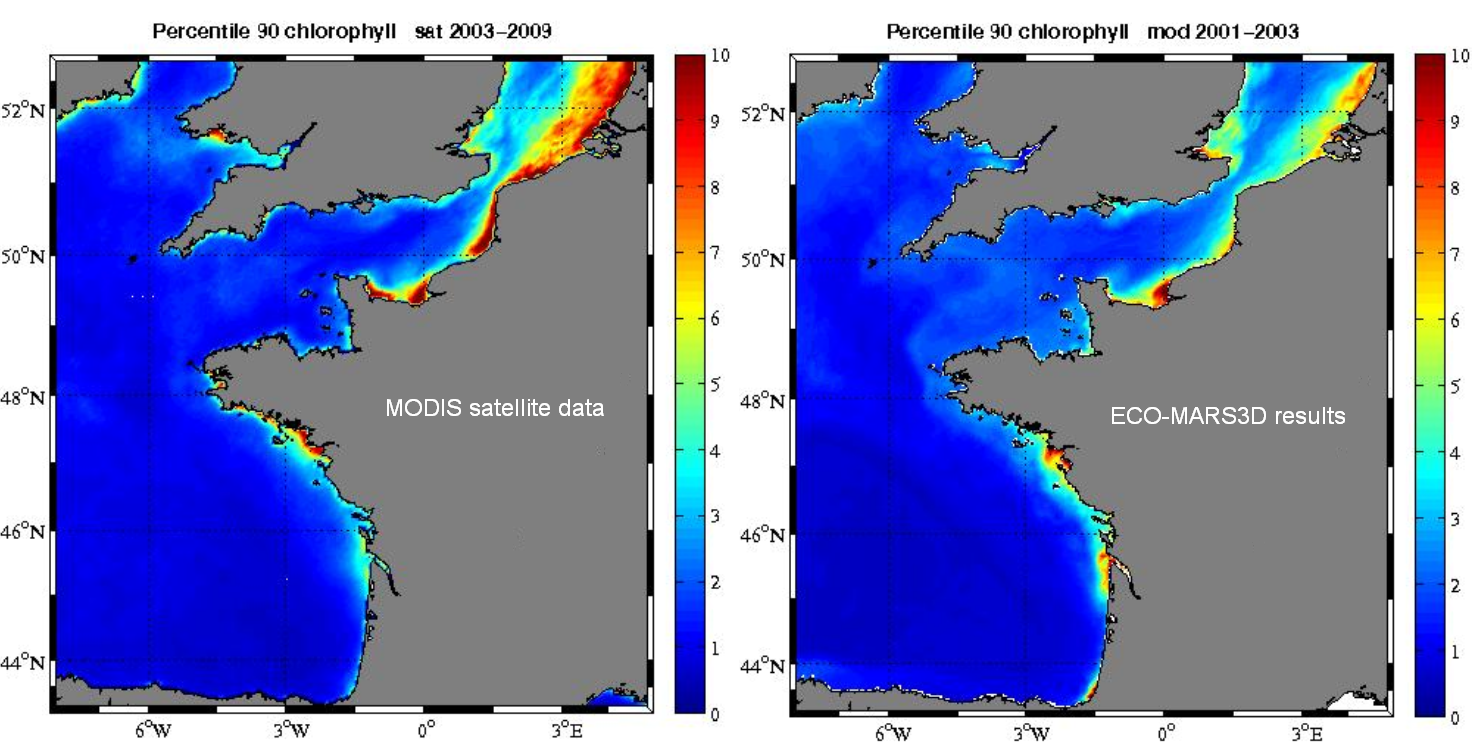
Figure 4. Validation of the simulated total chlorophyll by the corresponding satellite measurements (Dussauze, 2011)
The extended version of this biogeochemical model adds 3 harmful phytoplanktonic species (Pseudo-Nitzschia, Karenia, Phaeocystis) to the 3 bulk groups. Every specific phytoplanktonic module is derived from the corresponding bulk variable by replacing some ecophysiological parameters generic values by the values specifically related to the genus or species in the scientific literature, but also by modifying some process formulation, with possible adjunction of new state variables. For Pseudo-Nitzschia, the silicon mass is simulated independently from the basic nitrogen mass, which allows the simulation of the toxin (domoic acid) according to the Si/N intracellular quota (Davidson and Fehling, 2006). For Karenia, a daily vertical migration is added. For Phaeocystis, a simplified sub-model has been built from the original model of Lancelot et al. (2005); it uses only 3 state variables: the single motile cells, the colonies, and the mucus produced. For the three harmful species under study, the simple Q10=2 law for temperature effect on growth is replaced by a Gaussian curve, characterized by its mean (=optimal temperature) and its standard deviation.
The ECO-MARS3D model contains the add-on facility for nitrogen or phosphorus tracking (Ménesguen et al., 2006) (Fig. 5).
Figure 5. Nitrogen tracking in the food web: fraction of phytoplanktonic nitrogen coming from the Loire and Brittany rivers inputs. (Ménesguen, personal communication)
PCOMS
The Portuguese Coast Operational Modelling System (PCOMS) is based on the MOHID model operational platform for the West Iberian coast. The online one-way nested modelling system consists of two model configurations: a 2-D barotropic model with 0.06º of resolution forced only with the FES2004 tidal atlas solution using a Blumberg and Kantha (1985) radiation scheme and covering the geographic area 33.5° N - 45.9° N and 13.5° W - 4.2° W. The 2D model becomes the online external reference tidal solution to the west Iberia 3-D baroclinic model with the same horizontal resolution, covering the region 34.4° N - 45.0° N and 12.6° W - 5.5° W and with 50 vertical layers, where the bottom 43 are in Cartesian coordinates and the top 10 m are 7 sigma coordinate layers.
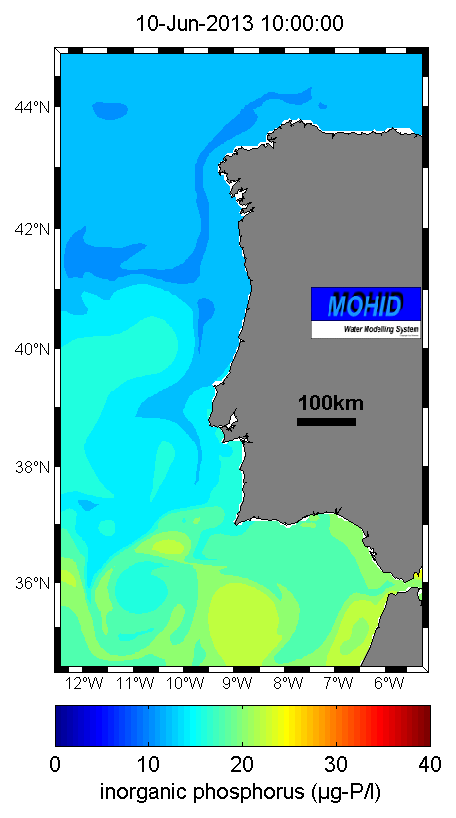
Figure 6. Inorganic phosporus forecast (10/06/2013) obtained with PCOMS (Source)
References
Davidson K., Fehling J., 2006. Modelling the influence of silicon and phosphorus limitation on the growth and toxicity of Pseudo-Nitzschia seriata. African Journal of Marine Science, 28(2), 357-360.
Dussauze M., 2011. Utilisation du modèle ECO-MARS 3D pour évaluer le bon état écologique de la façade Manche-Atlantique au regard du descripteur 5 "Eutrophisation" de la DCSMM - Phase 2. Ministère de l'Environnement, du Développement Durable, des Transports et du Logement, Ref. DCSMM 2011 Phase 1 – Rapport n POC.0399_1 - V2.0 – 08/11/2011, 45p.http://archimer.ifremer.fr/doc/00061/17206/
Gohin, F., Loyer, S., Lunven, M., Labry, C., Froidefond, J.-M., Delmas, D., Huret, M. and Herbland, A., 2005. Satellite-derived parameters for biological modelling in coastal waters: Illustration over the eastern continental shelf of the Bay of Biscay. Remote Sensing of Environment, 95: 29–46.
Guillaud, J.-F., L. Bouriel, 2007. Relationships between nitrate concentration and river flow, and temporal trends of nitrate in 25 rivers of Brittany (France). Revue des Sciences de l’Eau, 20(2) : 213-226.
Huret M., Sourisseau M., Petitgas P., Struski C., Léger F., Lazure P., 2013. A multi-decadal hindcast of a physical–biogeochemical model and derived oceanographic indices in the Bay of Biscay. Journal of Marine Systems, 109, S77-S94.
Lacroix G, Ruddick K, Park Y, Gypens N and Lancelot C (2007). Validation of the 3D biogeochemical model Miro&Co with field nutrient and phytoplankton data and Meris-derived surface chlorophyll a images. Journal of Marine Systems 64, 66-88.
Lancelot C, Spitz Y, Gypens N, Ruddick K, Becquevort S, Rousseau V, Lacroix G, Billen G (2005). Modelling diatom and Phaeocystis blooms and nutrient cycles in the Southern Bight of the North Sea: the MIRO model. Marine Ecology Progress Series 289, 63–78.
Lancelot C, Rousseau V, Gypens N (2009). Ecologicaly based indicators for Phaeocystis disturbance in eutrophied Belgian coastal waters (Southern North Sea) based on field observations and ecological modelling. Journal of Sea Research 61, 44-49.
Lazure P., Dumas F., 2008. An external–internal mode coupling for a 3D hydrodynamical model for applications at regional scale (MARS). Advances in Water Resources, 31(2), 233-250.
Luyten PJ, Jones JE, Proctor R, Tabor A, Tett P, Wild-Allen K (1999). COHERENS Documentation: a Coupled Hydrodynamical-Ecological Model for Regional and Shelf Seas: User Documentation. MUMM, Brussels.
Mateus, M., Riflet, G., Chambel, P., Fernandes, L., Fernandes, R., Juliano, M., Campuzano, F., et al. (2012). An operational model for the West Iberian coast: products and services. Ocean Science, 8, 713–732. doi:10.5194/os-8-713-2012.
Ménesguen, A., Cugier, P. and Leblond, I., 2006. A new numerical technique for tracking chemical species in a multisource, coastal ecosystem, applied to nitrogen causing Ulva blooms in the Bay of Brest (France). Limnology and Oceanography, 51(1, part 2): 591-601.